Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Kaloriferci Aday Tespit Listesi
gıda güvenliği uzman portalı
Analýza proliferace
GAZİOSMANPAŞA ÜNİVERSİTESİ
Show Full Text ()
Türkiye`de bağımsız bir hukuk bürosu olarak uluslararası hukuk ve
Termická analýza a kalorimetrie
DEFNE VE KEKİK YAĞI EKLENEN JELATİN İLE
linkten
Academic CVs - Çukurova Üniversitesi
Hijyen Nedir? - Erhan Ateş Kimya Okulu
EFFECTS of PREBIOTICS on GROWTH and
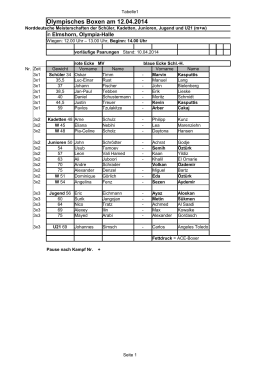
Olympisches Boxen am 12.04.2014
Hakem Onaylı Makale - Gıda Kontrol Laboratuvar Müdürlükleri
Sayı 8 - Mustafa Kemal Üniversitesi
Probiyotik Özellik Gösteren Bazı Laktik Asit
SET-VAC - Güneşli Aşı
S - Uzman Veteriner
setkání českých a německých spisovatelů na univerzitě hradec
DEKONTAMINASI BAKTERI Escherichia coli DAN Staphylococcus
Wchuwr/ …………………