Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Kıl folikülü kök hücreleri ve intrafoliküler homeostaz
Závěrečná zpráva 2013
Sayı: 3 - Kök Hücre E
KÖK HÜCRELER ve KLİNİKTE KULLANIMLARI STEM CELLS AND
FT Porte 1 vantail alu Baumert.indd
dp3 diyaframli pompa kullanma kilavuzu
Full Text Article - Hacettepe Üniversitesi
(PDF)... - DergiPark
Alt çeneye inokule olan cam parçalarının doğru
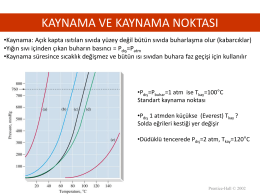
Ders 2 - fbuyukserin.at.etu.edu.tr
İçimizdeki Balık (okumak için tıkla)
Klasyfikacja i kryteria diagnostyczne nowotworów układu chłonnego
Devamı - kokhucrebulteni.com
Saç mikroskopisi ve trikogram
Epidermal Nevüsün Nadir Bir Varyantı: Bir Nevüs Komedonikus
Sağlık Olsun - Avcılar Hospital
Ad-Soyad: Süre: 40 dakika Sınıf-Numara: Not: Her soru 5 puan I
Czytadełko - Przedszkole 206
Anabilim Dalı Öğretim Üyesi Önerilen Tez Başlığı Anatomi Doç. Dr
Anti Anjijonik Tedavilerde Tartışmalı Konular
Czytadełko - Przedszkole 206
การศึกษาประสิทธิภาพการกระตุ้นการเติบโตของข
Slovník