Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
M Hücresi - Turkish Journal of Immunology
PROJEKT GILGAMEŠ
Tam metin için tıklayınız.
Ayrıntılı bilgi - Elektrik Tesisat Portalı
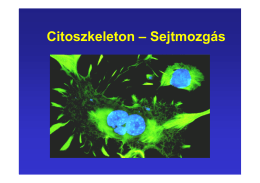
Citoszkeleton – Sejtmozgás
Immunoloji 2010:Layout 1 - Turkish Journal of Immunology
Download File
progresif familyal intrahepatik kolestaz
Ders 10 - fbuyukserin.at.etu.edu.tr
9 Biyoloji soru düşük çözünürlük PDF
Lézie tkaniva nervus vagus
ŞİLÖZ ASİT
The Study of the Methods and Types of Translating Onomatopoeia
Organigramme du Service d`immunologie et allergie, CHUV
cecf - program 2015.indd
Latina .pdf
Aphanius anatoliae sureyanus (Neu, 1937
KISA ÜRÜN BİLGİSİ 1. BEŞERİ TIBBİ ÜRÜNÜN ADI DROPOLEV 60
(PDF)... - DergiPark
Familial Mediterranean Fever Related Spondyloarthritis Ailesel
CEPE
Köpek ve Kedilerde Cerrahi Yara Enfeksiyonu ve Profilaktik
Kutay Biberoğlu