Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Genetic diversity of village chickens in Central Black Sea
Ders 6-Bilgi Arama-Veri Tabanlari
Bir Çok Boyutlu Ölçekleme Tekniği Olarak Torgersen Ölçekleme
error error error error error error error error error error error error
Územní rozhodnutí Villa Park ZTV
promoce2015 346KB Jun 02 2015 01:03:31 PM
AB Bölgesel Politikası Adalar ve Gökçeada
sajmovi u svetu 2015
dokumenta/propisi/pravila o radu distributivnog sistema Centar d.o.o
Tu bi, ili nije bilo tu
Állapotfelügyelet dokumentum
dekoratif tuğla teknik bülten
ระเบียบการใช้ห้องปฏิบัติการเคมีอาหาร
Référence : FE-3-010 (suite) Feuille d`exercices F - No
Horizonte 5/2014
Obor AMGP - Šlechtění zvířat
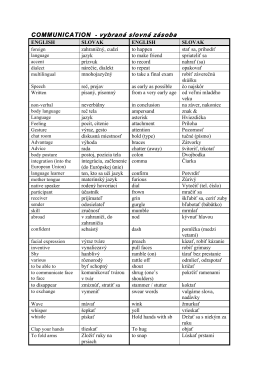
COMMUNICATION - vybraná slovná zásoba
Ime i prezime LEILA BEGIĆ Akademski naslov DOKTOR
ประสิทธิภาพการลงทุนที่วิเคราะห์โดยใช้ปัจจั P
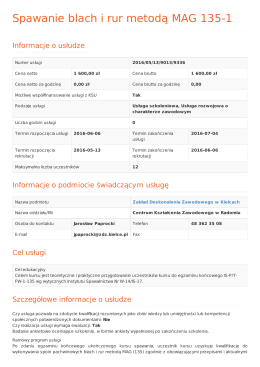
Spawanie blach i rur metodą MAG 135-1
ve Trichogramma euproctidis (Girault)
Press release HVS_PL
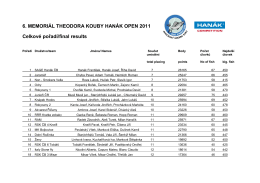
Koubův memoriál