Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
(cucurbita spp) species under drought stress
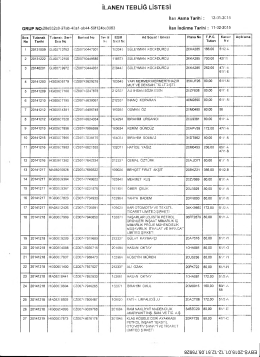
12.01.2015 Tarihli İlan Listesidir.
Düşük Büyüme, Orta Demokrasi, Yüksek Risk - Nisan 2014
Emniyet Kemeri Kullanımı 10.02.2015 14:26
Identity of Carrier klozu
Uchwala Nr VI/46/15 z dnia 23 lutego 2015 r.
CENÍK SLUŽEB
PSG Accessories 2013 Datasheet New Font - HP - Hewlett
Online monitor částic OPM II Typ OPM II
Zea mays, Poaceae - Biyoloji Kongreleri
spínaný napájací zdroj dc/dc pre priemyselné aplikácie
Effects of Interstock - Ankara Üniversitesi Dergiler Veritabanı
Sorular
Gülşah Altun
Obwieszczenie o okręgach wyborczych
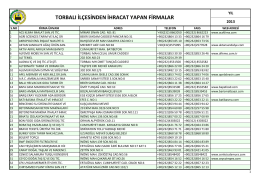
torbalı ilçesinden ihracat yapan firmalar
Bilinç ve Beyin : Bilincin Beyindeki Temsili
tc kafkas üniversitesi pedagojik formasyon sertifika programı a
the page - Balkan Agricultural Congress
Prezentacja dot. projektu Programu Strategicznego Włączenie
Düşük Doz Hidrojen Peroksit Ön MuamelesininKuraklık Toleransı