Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
14. Translasyon ve Proteinler.pptx
pdf format - Sanal Biyoloji
P12. Disiplinler Arası Bir Bilim Dalı: Biyoinformatik
Dumlupınar Üniversitesi Sosyal Bilimler Dergisi / Dumlupınar
çamşıhı, emlek ve ilbeyli yöresi âşıkları
Moleküler Onkolojik Testler ve Ci~Teknik Şartnamesi TESTİN ADI
eylül 2014 dönemi 2. deneme sınavı klinik bilimler testi soru ve
Veli Öğrenci Klavuzu
19. bölüm.pptx - Prof. Dr. Bektaş TEPE
Bu PDF dosyasını indir
Stiahnúť... - Rodenticid.sk
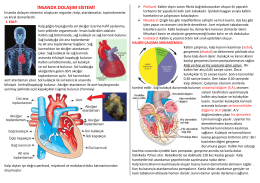
insanda dolaşım sistemi
BÖLÜM 12: Kromozomlardaki DNA`nın Organizasyonu
L I F E M A G A Z I N E
150-154 - türk biyokimya dergisi
ACTIVE DIRECTORY
Günlük Ağacı (Liquidambar orientalis, Sığla)
İnteraktif Oturum Tutanağı - Çevresel Gürültü Direktifinin Uygulama
daha fazla bilgi cv - Web ve Grafik Tasarım Uzmanı
2 - Zirve Üniversitesi
Product Leaflet: P-line 24 inç / 61 cm LCD monitör ve
Tıp_Ders3_Polar ve nötral amino asitler
Amino asitlerin sınıflandırılması