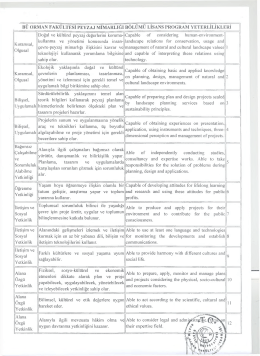
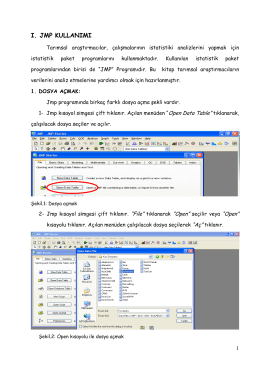
Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
ODRZIVI RAZVOJ.pdf
mini katalog izmene.cdr
stajnica knjiga.pmd
HANİFE TEZ 6.03.2014
A Guide for Enterprises “How can Enterprises support Vocational
mini katalog 012.cdr
Dosya İçin Tıklayın
Celé vydání
Vývoj finančních prostředků ve fondech EF
Cennik Oferty Internet Elastyczny 10.03.2016
Garantia en partes de diez (10) meses
John Grisham ÇINARLI YOL
Commercial Brochure (IP product overview)
Süt Sığırlarında Mevsimsel Beslemenin Sütün Karotenoid İçeriğine
E650 řady 3 (ZMD400AT/CT, ZFD400AT/CT)
değişik ekim ve dikim yöntemleriyle getirilmiş kızılcam
(MPPT)Tasarımı - Kocaeli Üniversitesi
tc sağlık bakanlığı türkiye halk sağlığı kurumu başkanlığı türkiye kalp
Dosyayı İndir
psikocoğrafya - WordPress.com
Cennik usługi Netia Mobile z komponentami