Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Zoológia bezchordátov
Zoológia 1
HAYVANLAR alemi - Biyoloji Bölümü
Spolupráca s praxou - Strojnícka fakulta
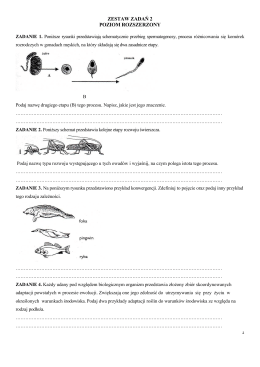
ZESTAW ZADAŃ 2 POZIOM ROZSZERZONY
ściągnij - JKK.edu.pl
Dynamiczne modele relacji interpersonalnych z opóšnieniem
TRITIKALE – Triticosecale / Wittm
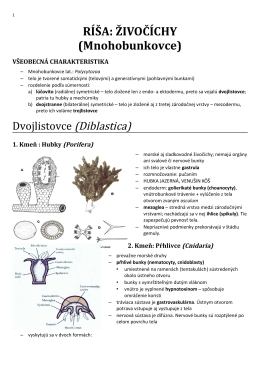
RÍŠA: ŽIVOČÍCHY (Mnohobunkovce)
Zadania - Korešpondenčný seminár z biológie
su omurgasızları laboratuvar kılavuzu
3 - Bohemia Cantat
Výkonový a honorářový řád a Unika pro Windows
ZOOLOGIJA INVERTEBRATA
Brwinów, dnia 2 grudnia 2013 r. Dyrektor Zespołu Szkół Nr 1 w
Phylum Porifera อ.แน็ต
Prehľad publikácií na predaj (Copy Centrum, Tajovského 10)
BAKALÁRSKE ŠTÚDIUM EKOLÓGIA 1. Bunkové základy
R 25
Poskytujeme komplexné riešenie vo svete obrábania
PROJEKTOVÁNÍ POZEMNÍCH KOMUNIKACÍ
Je biologická evolúcia slabo, stredne alebo silne usmerneným
katastrofální výsledek klinického testu objasněn