Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
zborník príspevkov konferencie centier excelentnosti
บทคัดย่อ - MBA Innovation Management
Kniha abstraktov - Intranet Katedry psychológie FiF UK
Scientific & social program - 13th Bratislava Symposium on
Zborník prispevkov a program - Institute Of Molecular Physiology
ZBORNÍK PRÍSPEVKOV 1. KONFERENCIE
Zarządzanie dużą infrastrukturą badawczą cz.II - Śląska Bio
sborníku 2012 - Ústav lékařské biochemie a laboratorní diagnostiky
Prof. Dr. N. Nuray Ulusu
Pokračovanie série článkov budúcich vedcov – chemikov
rebrové - as slovakia
9 - heidenhain
1. konferencia centra excelentnosti pre glykomiku
Lipopolysacharidy ako dominantné antigény bunkovej
RFID & NFC technologie pronikají do automobilového - smart-TEC
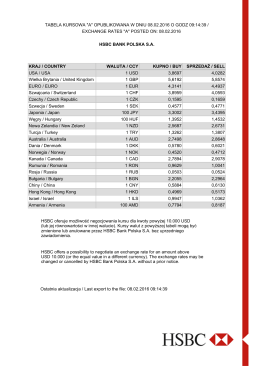
KRAJ / COUNTRY WALUTA / CCY KUPNO / BUY
REALBUD złączka zaciskowa z PP
Projekt Uchwały nr 3