Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
tek katlı zarla çevrili organeller
BİLİMSEL YÖNTEM - Biyoloji Bölümü
HAYVANLAR alemi - Biyoloji Bölümü
Miyokardın yapısı
Gelişim Psikolojisi Ek 1
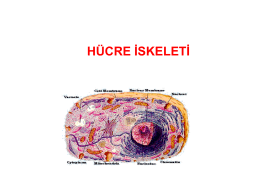
HÜCRE İSKELETİ
KISA ÜRÜN BİLGİSİ Bu ilaç ek izlemeye tabidir. Bu
XaXitest (4
TG – 4 - İhtiyaç Yayıncılık
Ders5
Hücre ve Hücre Organelleri
1. Osnovni topološki pojmovi u R U ovom predavanju skup realnih
Pederol Sprey
1-24 - zeynepkamiltipbulteni.org
Otoimmün büllü hastalıklar ve paraneoplazi
8. Eglenceli Okul Mart 2016 Yarım Günlük Plan
Tam Metin - Yaşam Bilimleri Dergisi
Kimya Mühendisliği Bölümü Bitirme Ödevi ve Lisans Araştırma
akılcı ilaç kullanımı - DOD Dermatoloji Gündemi
Biyoloji - İhtiyaç Akademi
PBC - Dynex
9 Biyoloji soru düşük çözünürlük PDF
T.C KARABÜK ÜNİVERSİTESİ Teknik Eğitim