Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Untitled - Genetická společnost Gregora Mendela
Hungarian Molecular Life Sciences 2013 Programme & Book of
XVII. konference - Fakulta veterinární hygieny a ekologie
STÁTNÍ ÚSTAV PRO KONTROLU LÉČIV
Genetika v biotechnologiích.pdf
pobierz program konferencji - e
ANCA ilişkili vaskülitler
Recenzja rozprawy doktorskiej pani mgr Katarzyny Anny Wójcik p.t.
oet 2 tehničari
obecná mikrobiologie (.pdf)
DİRENÇLİ GRAM POZİTİF KOKLAR: SAKIN KAÇIRMAYIN !
SPC
S. aureus
FTA karty - DYNEX.CZ
[Original]
Elektrotechnika 1
Łódź, dn. 08.09.2015r . Zapytanie ofertowe nr 1/OF/2015
Seniori
(mg/dl) séru v kreatinin x 72 (kg) hmotnost x (roky)] věk
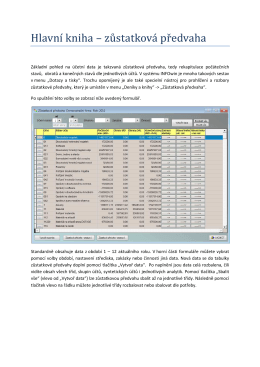
zůstatková předvaha
2016-04-15 TS ocenenie spolupracovnika UPJS LF z Holandska
Opracowanie nowego protokołu multiplexPCR
Na lekcjach języka polskiego












![[Original]](http://s2.readgur.com/store/data/000131121_1-b04390ce5ec73c1aed913e3639312962-260x520.png)



![(mg/dl) séru v kreatinin x 72 (kg) hmotnost x (roky)] věk](http://s1.readgur.com/store/data/000771475_1-dae75ed7c760d7f8b948b6a47b563815-260x520.png)