Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Vlastnosti půdy a struktura lesního ekosystému ve vztahu k
hıyar yetiştiriciliği
Vliv asanačních zásahů v oblasti Ptačího potoka (NP Šumava
(Coleoptera) mokrade Pramenisko pri Tatranskej Lomnici v Tatrách
KarmelInfo_62 - Karmelici Bosi
DivaDlem proti krizi
Dynamika lesa a krajiny jako podklad pro zonaci NP, aneb co
AGROSTEMIN
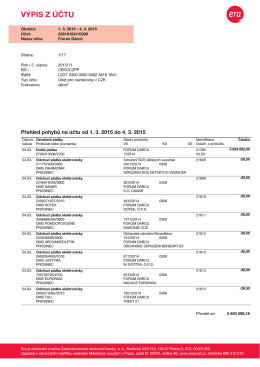
transparentním účtu
In memoriam Dr. Rajna Jovanović - Botanica Serbica
6569 SAYILI AF KANUNU senato kararı
Leden (.pdf) - Kulturní a komunitní centrum Přeštice
ArRevue 2014.pdf
2012 - Fakulta životního prostředí - Česká zemědělská univerzita v
Sestava 1 - Akademický bulletin
Václav, R. 2010. Súvisia reprodukčné parametre vrabca domového
wybór najkorzystniejszej oferty z dnia 03.07.2014
Zu Gast bei Freunden - Kinzig
Jak reaguje biodiver Jak reaguje biodiversita Šumavy na ita Šumavy
XI. Halászlé-TYR-kupa (Baja, 2013. november 30
Przeciwdziałanie bezpaństwowości dzieci: Analiza sytuacji w Polsce
Obrazowanie hiperspektralne – praktyczne
Pozvánka na závěrečný seminář