Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Proceedings SEED and SEEDLINGS X 2013
Poloprovozní pokusy kukuřice 2010
Vědecká příloha časopisu Úroda s příspěvky již ke stažení
Vědecká příloha časopisu Úroda s příspěvky již ke stažení
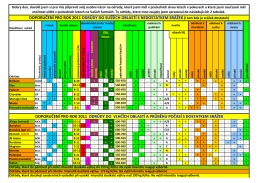
Odrůdová doporučení 2011 - pšenice ozimá
zde - EnergoAgro spol. s ro
Profesionálna ochrana rastlín
Duże wahania zbóż – teraz aa - Osadkowski
Eurogreen-katalog
Kniha ACTA 2007_1.indb - Acta Universitatis Agriculturae et
celý vestník - Ministerstvo pôdohospodárstva a rozvoja vidieka SR
HybRidní TRávníK - EUROGREEN
vliv abiotických a biotických stresorů na vlastnosti rostlin