Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Acta Facultatis Ecologiae
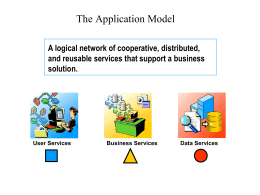
The Application Model
Identifikačný list nebezpečného odpadu
Jubilea členů ČBS v roce 2011 - Zprávy České botanické společnosti
Urbanistické středisko JIHLAVA, spol.s r.o.
Hornoorešan - číslo 1/2010
program konferencie
pošvatky (plecoptera) udavy (východné karpaty, slovensko)
tutaj - nasz lidzbark
Zadání
Sborník - Česká parazitologická společnost
Případová studie implementace EMC Avamar ve společnosti Teleplan.
Biyobenzer ürünlere genel bakış ve kılavuzlar
ČOV Prešov - KUNST, spol. s ro
zborník Zoológia 2012 - Conference system
Limnologické noviny č. 1/2012 - Česká limnologická společnost
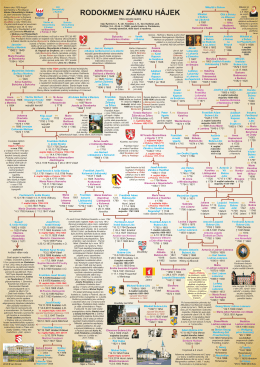
majitelů
Zpravodaj 24
szkolenia - Policja.pl
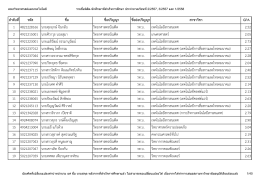
ลำดับที่ รหัส ชื่อ ชื่อปริญญำ ชื่อย่อปริญญำ สำขำวิชำ GPA