Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
6 Informační makromolekuly
W I A D O M O Ś C I K R A J O W E
ukázka
1.Ročník strukturovaného studia
REPLIKACE, TRANSKRIPCE, TRANSLACE - Bi
4 - Biochemie
Nevěřím, že by někdo byl tak naivní, aby si myslel, že
Genetika v biotechnologiích.pdf
1. Úvod
II.P.1 II.P.2 Session II. Regulation of gene expression in Prokaryotes
książkę abstraktów - Sopockie Towarzystwo Naukowe
Poštovní podmínky Ostatní služby
(Microsoft PowerPoint - P1 LS - Z\341klady biochemie a metabolismu)
NMR - lekce9
I rostliny mají hormony [pdf]
BÖLÜM 12: Kromozomlardaki DNA`nın Organizasyonu
Franz Boas: Lingvistika a etnologie (pdf)
Organické látky - transformace a degradace
Úvod do chemie léčiv
Základní parametry absorpčního spektra, vliv přístrojové funkce
Porovnani ucinku mleti na strukturu a vlastnosti mouk
Stanovení obsahu alkoholu ve vzorku piva
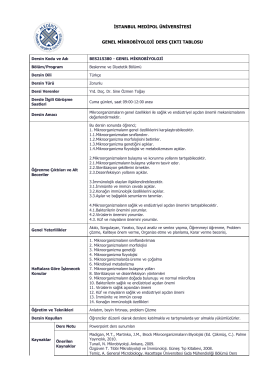
istanbul medipol üniversitesi genel mikrobiyoloji ders çıktı tablosu











![I rostliny mají hormony [pdf]](http://s2.readgur.com/store/data/000152782_1-44a40b4f530386dabc9821997c460986-260x520.png)