Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Adaptace-Potrava
5. přednáška
PDF verze odborného programu - Oční klinika Horní Počernice
Mammalogie
Makaleyi İndir - Veteriner Kontrol Enstitüsü Müdürlükleri
Biologická antropologie
otevřít
Výše úhrad a podmínky úhrady ambulantních hrazených
Cricetinae and Murinae (Rodentia ) from Bacho Kiro Cave, Bulgaria
Fyziologie II – 2016/2017
Fyziologie II – 2015/2016
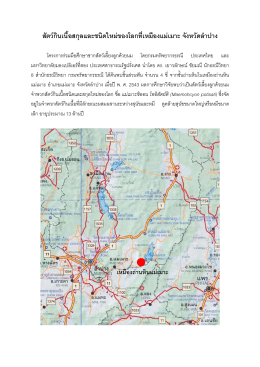
สัตว กินเนื้อสกุลและชนิดใหม ของโลกที่เหมือ
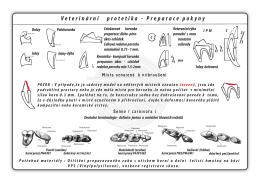
Veterinární protetika preparace pokyny
11. Amniota: Lepidosauria
Mammalia - ČR
Program - Krajská zdravotní, as
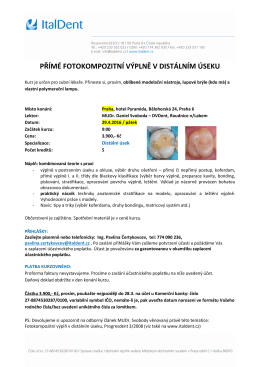
zde - ItalDent
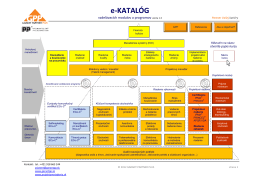
e-KATALÓG - Projektové riadenie