Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
line - Biotechnologická společnost
Chemická sekcia
PDF İndir - Türkiye Aile Hekimliği Dergisi
Lista zabranjenih sredstava i metoda u 2010 godini
první linie_2_2012
Synlabianer 04/2014 - synlab Czech Republic
sborník přednášek a posterů
Draft TOC for ETFA06, Mode warn, Ver
Chemie - 010 - Organická chemie
Dr. Max
prospect bi - Biotechnologická společnost
Bioprospect_1_11.qxd:Layout 1
TYLNA STRONA - NAKLEJKA PRZECIW PLEŚNI
HISTORIE IMUNOLOGIE
Dystrybutor: PROX Sp. z o.o., ul. Parkowa 25, 51
vyšetrovanie mechanických vlastností materiálov metódou
IDE - SecuriFocus
pada 2009 r. o grach hazardowych (Dz. U. Nr 201, poz. 1540, z 2
BULLETIN BIOTECHNOLOGICKÉ SPOLEČNOSTI zakládajícího
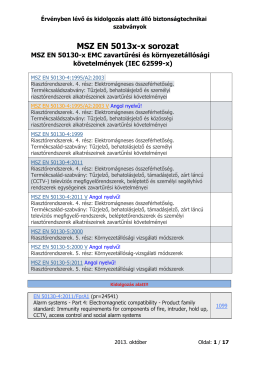
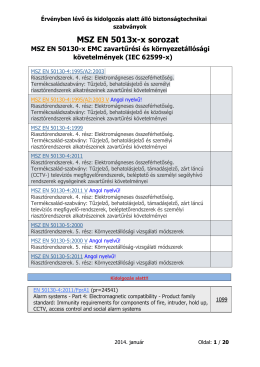
Érvényben lévő és kidolgozás alatt álló biztonságtechnikai
novém čísle Bioprospektu (4/2015)
letöltés