Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
12 Základy virologie
Chřipka a těhotenství
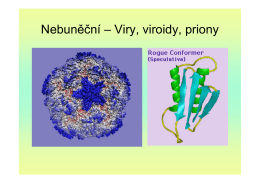
Nebuněční – Viry, viroidy, priony
8. Viry
Mgr. Michal Křupka
Biotechnologický ústav AV ČR, vvi
AP-24® - Nu Skin
Nöroleptik malign sendrom tablosunda acil
LABORATORNÍ LISTY LABORATORNÍ LISTY
Zadání
Enfeksiyöz Bronşitisli Broylerlerde IB Virus Genotipleri: Ülkemizdeki
Zdraví národa vstupuje do ordinací i k pacientům Jak čelit tuberkulóze
Diagnostika rostlinných virů - Biologické centrum AV ČR, vvi
zobrazit
4. Genové inženýrství ve farmaceutické biotechnologii
HISTORIE IMUNOLOGIE
učebný text - Gymnázium Štefana Moysesa MOLDAVA NAD BODVOU
EXCELLYSE Live 100 ml EXCELLYSE Live 30 ml
OKNA DVEŘE VRATA
Wyciszanie genów zapobiega chorobie wściekłych
SCZ7100 – BIPV fasáda
Výroční zpráva 2015 - 22. středisko Junáka Brno
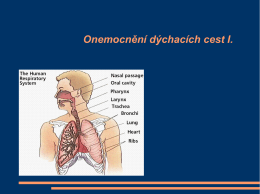
Onemocnění dýchacích cest I .