Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
PROSPECT B I - Biotechnologická společnost
Stručný průvodce systémem Windows 8 Cloudové
Aký je benefit pre matku a dieťa z prenatálnych skríningov a
sešit 4 - Ksicht
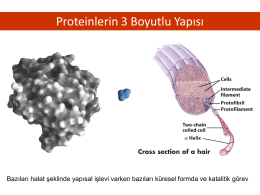
Proteinlerin 3 Boyutlu Yapısı
ZDE - Laboratoř pro vývoj a realizaci
Klonování a sekvenování přírodní DNA
Článek v PDF ke stažení
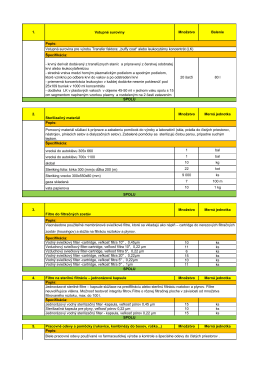
1. Vstupné suroviny Množstvo Balenie
Diferenciální diagnostika bolestí hlavy.
doa keluarga - Gereja Santo Yakobus , Paroki Kelapa Gading, Jakarta
Studijní text - Biologická olympiáda
Otázky ke zkoušce
Demence - AKTUÁLNĚ
null
Ryby našich vod
Otiskovací hmoty 2012
Sınıf Öğretmenlerinin Öğrencilerle İletişim Becerileri[1 - CYBD-JCAS
Kapitula 5/2014
Případ č. 26
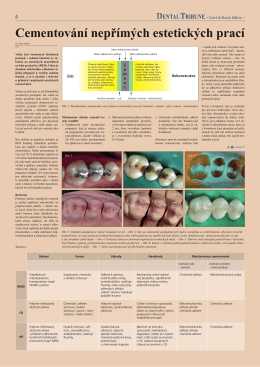
Cementování nepřímých estetických prací
Zprávy - Chemické listy
Doporučené postupy - Klinická onkologie