Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
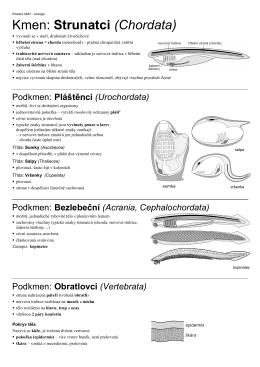
CHORDATA
4. Tunicata - Webnode.cz
4. Craniata - Obratlovci
Subdivisio: Enterocoela
prezentace
Obecná zoologie - KATEDRA BIOLOGIE
Dobra praktyka nadzoru weterynaryjnego w zakresie dobrostanu ryb
Презентација 01 Вода и адаптације
Otázky, na které kreacionismus nemá odpověď
Květen
biologija
ความหลากหลายทางชีวภาพของเพรียงหัวหอม ในแนวป
14.9.2015 1 Pláštěnci – Urochordata = Tunicata Tunicata v systému
1. Koacerváty vznikali (označte správnu odpoveď): A
Avis de mise en candidature - Conseil des écoles fransaskoises
Přehled GMH Strunatci
Zobrazovací diagnostika tumorů páteře
Profilové zkoušky MZ
Podkmen: Pláštěnci
Seznam publikované literatury
Přehledová práce Fetální MR CNS u poruch ventrální
Životopis a odborný profil
Technický list