Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
zde - Restoration Ecology Group
LP-CIGP
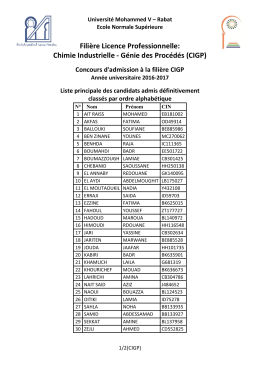
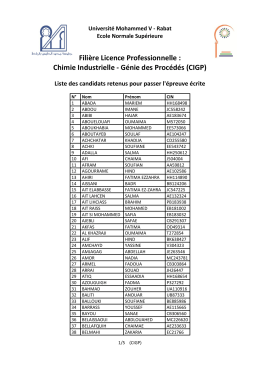
Filière Licence Professionnelle : Chimie Industrielle
Klaňte se spontánní sukcesi! …protože přátelé… ona funguje!!!
Zdislavská pouť 2014
ŠVNS ZŠ a MŠ České Velenice
Výroční zpráva - Švehlova střední škola Prostějov
Summary news (สรุปข่าวประจำสัปดาห์)
Ochrana přírody 2.indd
Umowa BZWBK24-B-OS.FIZYCZNA
Katalog poskytovatelů sociálních a doprovodných služeb na
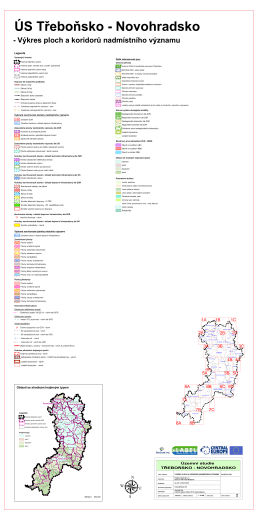
- Výkres ploch a koridorů nadmístního významu
Dossier_de_presse_A75_Issoire-final
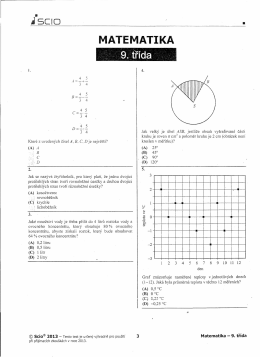
Scio test 2014 - Vasekpasek.cz
Program 19. SHMO ČR
eGovernmt, PDF - IT Asociácia Slovenska
Calendrier des épreuves écrites - Université Mohammed V de Rabat
Erik Satie - Gnossienne No. 3