Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
TRAKYA ÜNĠVERSĠTESĠ FEN BĠLĠMLERĠ ENSTĠTÜSÜ
FENOLİK BİLEŞİKLERİN TAYİNİNE YÖNELİK AMPEROMETRİK
T.C. NEVġEHĠR HACI BEKTAġ VELĠ ÜNĠVERSĠTESĠ
bölüm
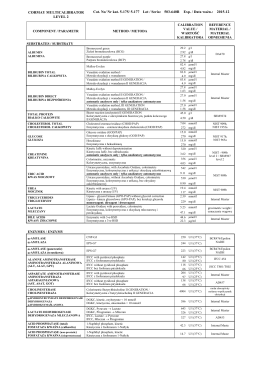
CORMAY SERUM HN
Uluslararası İnsan Hakları Belgelerinde Sosyal Koruma Hakkı
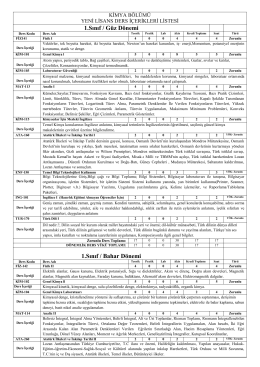
Kimya Bölümü Ders İçerikleri
T.C. GAZİOSMANPAŞA ÜNİVERSİTESİ
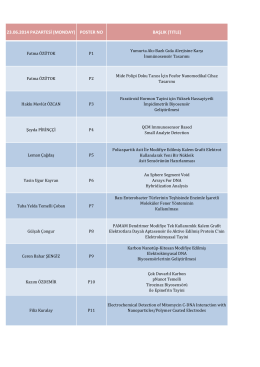
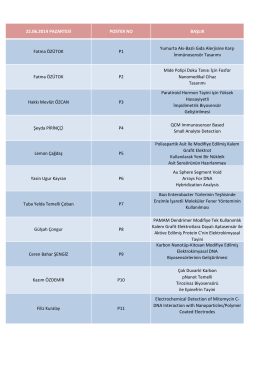
23.06.2014 pazartesi (monday) poster no başlık
TIPTA UZMANLIK EĞİTİMİ GİRİŞ SINAVI (TUS) (Sonbahar Dönemi
Yaşamın Kökeni, Evrim ve Tanrı - Ankara Üniversitesi Dergiler
STERILIZASYON U NITESI ISLEYIS PROSEDÜRÜ
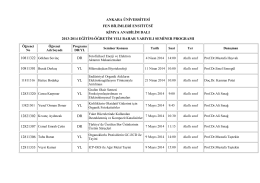
ankara üniversitesi fen bilimleri enstitüsü kimya anabilim dalı 2013
DASRehber 15ocak2014-
www.adanabtu.edu.tr - Biyomühendislik
22.06.2014 Pazar (Sunday) - E
Fen Bilimleri Enstitüsü
4(2): 54-58 Altay ve ark. 54 Original Article / Orijinal Araştırma
22.06.2014 PAZARTESİ POSTER NO BAŞLIK Fatma ÖZÜTOK P1
Mühendislik Mimarlık Fakültesi
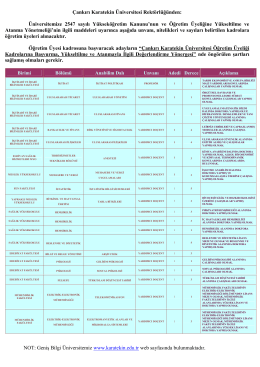
Çankırı Karatekin Üniversitesi Rektörlüğünden: Üniversitemize 2547
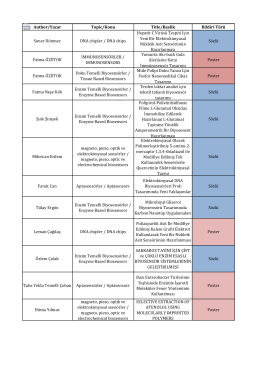
Author/Yazar Topic/Konu Title/Baslik Bildiri Türü Soner Dönmez
tc manġsa ġlġ halk sağlığı klġnġk laboratuarı test rehberġ