Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
05 Meióza
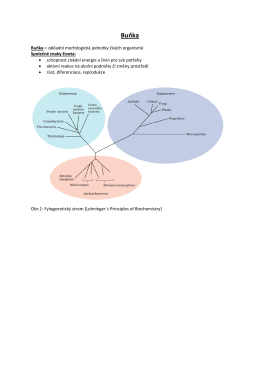
Buňka - Wiki Matfyz
Buněčný cyklus Přípravný kurz
04 Mitóza a buněčný cyklus
Genomová nestabilita kmenových buněk
Sarkoid u - Veterinární klinika Doksy
Co je to Pervitin? - P
SZZ - okruhy - Katedra ekologie a životního prostředí
Návod k obsluze IQ displej
stáhnout - IMPROMAT KLIMA
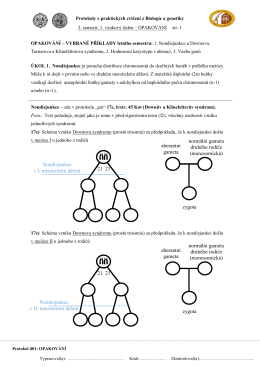
Morganove pravidlá
MECHANISMY EVOLUCE
Přírodovědný KLOKAN 2013/2014
podzim pití final
Buněčný cyklus Buňky žijí od dělení k dělení. Celý jejich život
Evoluční tání aneb o původu rodů
Ing. Antonín Fojtík, CSc. šlechtitel, genetik, pedagog
1. výukový týden
MUR-05
Nastavení pro propojení se SW Pohoda
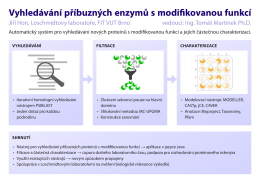
Vyhledávání příbuzných enzymů s modifikovanou funkcí
PDF | Lymfomy GIT