Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
Rajtova teorija pomične Rajtova teorija pomične ravnoteže
i P I S M O - masterlogo.pl
I . III . II I. YARIYIL III. YARIYIL I . III . IIK ü l K ü l Kültür ve İstanbul
Müdürlüğümüzün konu ile ilgili 09/04/2014 tarih ve 1461293 sayılı
Leteća pešadija.PDF
Murat Vural v. Türkiye - İnsan Hakları Daire Başkanlığı
slobodna misao iakcija
วาระการประชุม
preuzmite brošuru - Evropski Dan Srčane Slabosti
prirodne organske materije u vodi – osvrt na
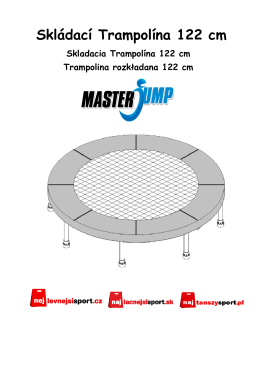
Návod na malé skládací trampolíny MASTERJUMP
Esemestník 3/2 - Slovenská mineralogická spoločnosť
TG – 3 - İhtiyaç Yayıncılık
Ogłoszenie o zamówieniu - BIP
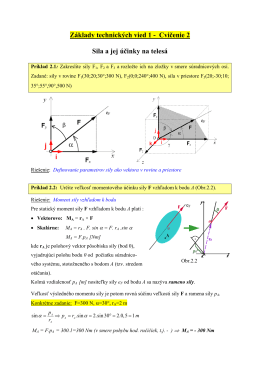
Základy technických vied 1 - Cvičenie 2 Sila a jej účinky na telesá
1. Oblicz częstość q występowania allelu a w populacji, w której 90
ภาพนิ่ง 1 - GEOCITIES.ws