Read
Gur
☰
Explore
Log in
Create new account
Upload
×
Download
No category
TEORIJA ORGANSKE EVOLUCIJE
Pogledaj PDF
Online Katalog
Mehanika stena i tla
животиње-кратак курс 2
knjiga sažetaka књига сажетака - Društvo psihologa Brčko Distrikta
SMOLUĆA
A vak órásmester
SPECIJACIJA. Č GENETIČKE DISTANCE
DN-B-I6 Morfometrija
interaktif su oyun grupları
Çözüm
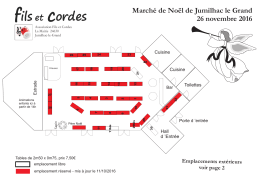
Plan de la salle
PP lepiace pásky s potlačou
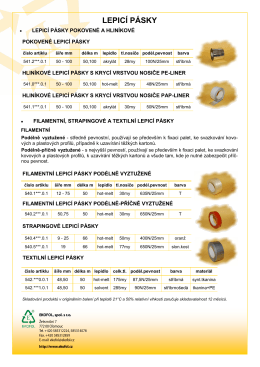
LEPICÍ PÁSKY
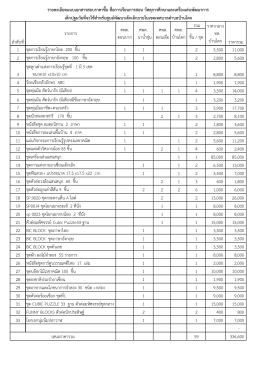
รวม ชิ้น / ชุด 1 ชุดการเรียนรู้ภาษาไทย 200 ชิ้น 1 1 2 5